


ULTRASTRUCTURE AND HISTOCHEMISTRY OF ANTHER DEVELOPMENT IN MELIA AZEDARACH L.
HTML Full TextULTRASTRUCTURE AND HISTOCHEMISTRY OF ANTHER DEVELOPMENT IN MELIA AZEDARACH L.
P. L. Talawar * and S. N. Agadi
Developmental Biology and Histochemistry Laboratory, P. G. Department of Botany, Karnatak University, Dharwad - 580003, Karnataka, India.
ABSTRACT: We provide evidence of the structural development and distributional pattern of polysaccharides of the anthers in Melia azedarach L. The anthers of Melia azedarach L. is tetrasporangiate and basic type of wall formation is seen, it consists of six wall layers i.e. single layer of epidermis and endothecium, three middle wall layers and one tapetal layer. The primordium consists of a homogenous mass of meristematic cells which has a uniform distribution of insoluble polysaccharides. Sporogenous cells are moderate with PAS-positive and starch grains are present in all the wall layers. Meiocyte shows a moderate amount of carbohydrate with a prominent nucleus. The secretory cells are present near the connective and tapetum. Secretary cells are having abundant starch grains. Tetrahedral tetrads are present with surrounding thick callose walls with rich PAS-positive walls. The secretary type of tapetum is initially single-layered but some of the cells divide periclinally so that it becomes two-layered at vacuolated microspore stage. Middle wall layers are starting to degenerate at the tetrad stage. The pollen grains are tetracolporate and it contains abundant starch grains. The multiple fibrous thickening is formed in the dehisced stage with the rich content of polysaccharides. Dehiscence takes place through longitudinal slits and one middle wall layer is persistent at the dehisced stage.
Keywords: Meiocyte, Callose, Polysaccharides, Fibrous thickening, Tetrad
INTRODUCTION: Higher plants have a complex life cycle that alternates between the growth of a diploid sporophytic organism and highly reduced haploid gametophytic form. In flowering plants, the male gametophyte (or pollen grain) plays a vital role in plant fertility and crop production through the generation and delivery of the male gametes to the embryo sac for double fertilization.
During the reproductive stage, cells communicate closely with each other and as a result, male and female gametes are established independently 1, 2. The life cycle of flowering plants involves the alternations of a diploid sporophytic and haploid gametophytic phase 3. Microsporogenesis occurs at the end of the sporophytic phase and undergoes a clearly defined cell division program resulting in a gametophyte (pollen grain) comprising the generative and vegetative cells. The generative cell divides giving rise to two sperm cells 4.
Male gametophyte development occurs in four phases which differ in relation to environment and hydration state. 1. Pollen develops inside the anther with grains immersed in locular fluid which conveys substances from the mother plant. 2. The locular fluid disappears by reabsorption and/ or evaporation before the anther opens 5. 3. The anther opens and pollen may leave the anther as soon as it opens, or wait to be dispersed being glued with pollen kit, as occurs in almost all entomophilous species 6. 4. Pollen is dispersed by different agents remaining exposed for different periods.
The diversity of cells and tissues, developmental events and critical stages during anther ontogenesis has been at the center of interest since generative reproduction in plants depends on proper pollen structure and function. The chance of effective pollination and fertilization of maternal plants decreases due to defects in the development of the microspore or the surrounding nutritive layer, the tapetum 7. In addition, pollen coat components are secreted from active tapetal cells, which undergo structural and biochemical changes during the final phase of cell differentiation and death 8.
Histochemical assessment using a light microscope is of specific advantage, in getting a large quantity of data regarding the biochemical basis of tissue and organ differentiation such as anther. Histochemistry is concerned with the identification and distribution of various chemical components of tissues through the use of stains, indicators as well as microscopy. Histochemical studies have revealed the major biochemical substance i.e. carbohydrate plays an important role in the development and differentiation of various structure in the anther. Taylor and Osborn (2006) 9 stated that investigations of pollen structure and development and the accompanying changes within the anther yield an array of characters potentially useful for assessing phylogenetic relationships.
The Meliaceae family is having a number of medicinal plants whereas Azadirachta indica, Melia azadirachta, Khaya senegalensis, Swietenia mahogany etc. Various species are used for vegetable oil, soap – making, insecticides, and highly prized wood (mahogany). The Melia azedarach L. plant is commonly grown in Karnataka especially seen in the Dharwad region. Previous studies on this species focused on taxonomic features, Pharmacology, Tissue culture, Morphology etc. Literature about the embryology in species of the family Meliaceae is scanty and there is no recent work on this subject 10. In Melia azedarach, the development of the megagametophyte was described as Polygonum type 11. Galati et al., (2011) 12 worked in embryological studies in Melia azedarach L. Chang and Wang (1956) 13 studied the morphology of pollen grains in some members of the Meliaceae family. Garudamma (1956) 14 investigated embryogeny in Azadirachta indica A. Juss.
In Melia azedarach L. flowers, anthers, and stigma are positioned close to each other makes it possible for self-pollination. Eight yellow and hairy anthers were found surrounding the stigma in each flower 15. After surveying the literature we came to know that there is no anther development and histochemistry work has been done on Melia azedarach L. Talawar P. L. and Agadi S. N. (2018) 16 have studied the anther development and its polysaccharides distribution of Azadirachta indica A. Juss., now it’s time to focusing on other species of Meliaceae members. That’s why the present investigation was carried out to study the anther development and its distributional pattern of polysaccharide in Melia azedarach L. species. The present investigation gives a comprehensive account of the anatomical and histochemical changes during anther development.
The present study includes different developmental stages and insoluble polysaccharides distribution analysis of Melia azedarach L. anther with the following objectives.
- To study the different stages of anther development in Melia azedarach
- To understand the insoluble polysaccharides distribution in developing anthers of Melia azedarach
MATERIALS AND METHODS: The flower buds of Melia azedarach L. were collected from the University of Agricultural Science, Dharwad. The flower buds are fixed in Formalin –Acetic acid- Alcohol (FAA) for 12 h. Following fixation, materials were processed using conventional microtechnique methods of dehydration and infiltration using alcohol butanol series and embedded in paraffin wax. Sections were taken at 6µm thickness with Leica rotary microtome.
In the present investigation, Periodic Acid Schiff’s (PAS) method is employed to localize the total insoluble polysaccharides.
Total Insoluble Polysaccharide; Periodic Acid Schiff’s Method: 17 Periodic Acid Schiff’s (PAS) is staining is mainly used for staining structures containing a high proportion of carbohydrate macromolecules. PAS stain used in combination with diastase, an enzyme that breaks down glycogen. The reaction in the stain involves periodic acting as an oxidizing agent to oxidize the carbon – to – carbon bonds between two adjacent hydroxyl groups. This produces Schiff’s reactive aldehyde groups. The Schiff’s reagent comprises a mixture of basic fuchsin in the mixture reacts with newly formed aldehyde groups in the tissue to produce a bright magenta color.
Staining Procedure:
- Sections were deparaffinized in Pure Xylene for 30 min.
- Deparaffinized slides were treated with Alcohol: Xylene series i.e. (1:3, 1:1; 3:1) for two min in each series.
- Incubate the slides to alcohol series (100% to 50%) for two minutes in each series.
- Now slides were transferred to tap water for 5 minutes.
- The slides were incubated in 0.5% periodic acid for 15 min at room temperature and then rinsed with tap water.
- Now slides were treated with Schiff's reagent for 30 min at room temperature.
- Rinse the slides in Tap water for 5 min.
- Then rinse the slides in distilled water.
- Dehydration process with slides to alcohol series from (50%-100%) 2 min in each.
- Then slides were transferred to Alcohol: Xylene series (3:1, 1:1, 1:3) 2 minutes in each series.
- Finally, the slides are incubated in pure Xylene for 2 min.
- Mount the slides in DPX and observe the slides under a microscope for localization.
Colour Indication: Carbohydrate appears magenta-red color.
Photomicrography: The stained sections were microphotographed by using Samsung Galaxy A7 mobile 24 Megapixel camera with Olympus bright field microscope. The Schiff’s reagent is to produce magenta-red color. The assessment of total polysaccharides in the cells and tissues is qualitative. The results presented here are principally based on the intensity of the color in the cytoplasm and nuclei normal, rich and intense.
Observation: Anther development in plants is a systematic and complex process. Microsporo-genesis is the process in which each microspore mother cell divides meiotically to form four haploid microspores or pollen. The anther of Melia azedarach L. is tetrasporangiate. Anther develops from a mass of undifferentiated primordial cells to a complex set of tissues with varying functions.
A very young anther is oval shape and initially comprises a mass of undifferentiated meristematic cells surrounded by a well defined single layer of the epidermis. As development proceeds, the oval shape primordial becomes four-lobed anthers. The development of pollen sacs begins with the differentiation of archesporial cells in the hypodermal region below the epidermis at four corners of the young anther. The archesporial cells differ from other cells and are larger and contain dense cytoplasm with the prominent nucleus at the centre. This cell undergoes the first periclinal division to give a subepidermal primary parietal cell and inner primary sporogenous cell.
Further, the primary parietal cell undergoes periclinal division to produce outer secondary parietal cells and inner secondary parietal cells. The outer secondary parietal cell will undergo periclinal division and give rise to outer endothecium and inner middle wall layer. The middle wall layer is again divided by periclinal to give rise to another middle layer. Then the inner secondary parietal cells undergo the periclinal and to form one more middle wall layer outside and innermost tapetal layer.
Finally, the anthers having the single layered epidermis, a single-layered endothecium, three middle wall layers and a single-layered tapetum and conforms to the basic type of anther wall development. After the formation of the anther wall, primary sporogenous cells undergo repeated mitotic divisions and form a mass of sporogenous tissue. The epidermis is the outermost single layer and it is compactly arranged and usually protective in function. Tapetum is the innermost layer of anther walls and it completely surrounds the sporogenous tissue. The secretary type of tapetum is seen in Melia azedarach L. Tapetum is contains dense cytoplasm with prominent nuclei and it transports the nutrients to the developing sporocytes.
The cells of sporogenous tissue are polygonal in the outline and attached close to each other within the locule. The cells of sporogenous tissue have dense cytoplasm with the prominent nucleus. In the early sporogenous stage, the inner walls are thinner and later there is a deposition of the callose wall around each cell to get the inner wall of the cells to become thick-walled. The rich starch grains are present in all the wall layers in the beginning. The tapetum layer of developing anther surrounds the sporogenous tissue and plays an important role in the nutrition of microspores.
Later on, sporogenous cells give rise to microspore mother cells. As development proceeds the tapetal layer shows a lack of starch grains and secretary cells are present in connective cells and these cells are consisting of reserve food materials.
There are two ephemeral middle wall layers and one tapetal layer is present. During the onset of meiosis in the microspore mother cells a special callose wall layer is secreted around each microspore mother cell. The tapetum is intact at this stage. The microspore mother cells undergo Ist meiotic division and a binucleate cell is formed without wall formation. Hence, dyads are absent. So the meiosis in a microspore mother cell is accompanied by simultaneous cytokinesis. Later the two haploid nuclei undergo the second meiotic division giving rise to the tetrad. The tetrads were arranged in a tetrahedral manner. Each tetrahedral tetrad was surrounded by a thick callose envelope. The microspores within tetrad have dense cytoplasm with the prominent nucleus. One middle layer is starting to degenerate at this stage.
Later on, the callose wall is degraded by the activity of an enzyme callase secreted by the tapetum leading to the separation of tetrads into individual young microspores. The young microspores are spherical in shape and single-layered with the prominent nucleus. Tapetum is intact at this stage.
Later the young microspores are enlarged in its size and become vacuolated. In the vacuolated microspores, the protoplasm move to the periphery and also the nucleus displaced to a peripheral position in the thin layer of the cytoplasm. The tapetum is usually single-layered but some of the cells divide periclinally so that it becomes two-layered at this stage. The tapetal cells are at first uninucleate but due to mitotic divisions they become binucleate and each cell contains 1 to 2 nuclei. At this stage, the starch grains are only restricted to endothelial cells and when development occurs, the starch grains are completely disappearing in the mature anther. Vacuolated microspores accumulate starch grains and become young pollen grains. The young pollen grain has a single-layered wall, dense cytoplasm and a centrally situated single haploid nucleus. After sometimes the wall gets divided into two layers the outer exine and inner intine. The intine is thin and smooth but the exine is thick. The cells of the endothelium are radially elongated and show fibrous bands. The fibrous band's thickenings are made up of callose arise from the inner tangential walls.
The tapetum and one middle wall layer is intact at this stage. The epidermis shows papillate at this stage. The epidermis and endothecium of anther wall remain intact until the end of anther and pollen development. The first division of pollen grains gives rise to small generative cell and large vegetative cell. Endothelial cells help in the dehiscence of anther at maturity. The mature anther shows papillate epidermis, fibrous band thickenings and completely degenerated tapetum. One middle wall layer is persistent at the dehisced stage. The dehiscence is longitudinal in nature and mature pollen grains are scattered to the environment concurrent with dehiscence of stomium.
Distributional Pattern of Polysaccharide in the Anthers of Melia azedarach L: During the anther development, the distribution of insoluble poly-saccharides in the anther correlates with the different stages of male gametophyte development.
In developing anther tissues, insoluble polysaccharides are found in the form of PAS-positive tinge in the cytoplasm, starch storage and cell thickenings 18.
In Melia azedarach L. the transverse section of anther primordial meristematic cells having dense cytoplasm is enveloped by a well defined layer of the epidermis. The primordium consists of thin-walled cells distinctly PAS-positive having a uniform distribution of insoluble polysaccharides Fig. 1. In the sporogenous cell stage, before the anther wall completes its differentiation, cells of primary parietal layer, secondary parietal layer and primary sporogenous cells are rich in insoluble polysaccharides with abundant starch grains Fig. 2. Epidermis and endothecium are having rich PAS-positive. The tapetum and sporogenous cells are containing a moderate amount of polysaccharides. Periodic acid- Schiff's (PAS) staining showed that a large number of PAS-positive starch granules were present in wall layers at the early stage. The secretory cells are present in connective cells and consist of abundant reserve food materials which are highly reacted with PAS-positive reaction Fig. 3. At a late sporogenous stage, the wall layers are clearly seen i.e. single layer of epidermis and endothecium, three middle wall layers and single tapetum layer. Sporogenous tissues are located at the center and are moderately stained with polysaccharides. Accumulation of large amounts of PAS-positive granules in the anther wall layers Fig. 4. During the developmental stage, the polysaccharides distribution is increasing in the wall layers except for tapetum. Meiocytes are moderately stained and starch storage is more in meiocyte Fig. 5. In the following tetrad stage, the tetrahedral tetrads are enveloped by a richly stained callose wall. There is an increase in the number of polysaccharides in the anther wall layers except tapetum and high level of starch grains Fig. 6.

PLATE 1: TRANSVERSE SECTION OF MELIA AZEDARACH L. ANTHERS TESTED FOR INSOLUBLE POLYSACCHARIDES. (EP- Epidermis; End-Endothecium; WL- Wall layers; Mc- Mass of cells; PPC- Primary parietal cell; SPC – Sporogenous parietal cell; PSP- Primary sporogenous; Sp- Sporogenous; M- Meiocytes; T- Tapetum; Td-Tetrad; STG – Starch grain; Sc- Secretary cell)
- S. of anther primordia shows a mass of cells and epidermis with uniform distribution of polysaccharides.
- The archesporial cells show division and form primary parietal cells, secondary parietal and primary sporogenous cells. These cells are rich PAS +ve and abundant starch grains are present.
- At a sporogenous stage, the epidermis, endothecium and secretory cells show rich polysaccharides compared to sporogenous and tapetum. Starch grains are present in all the wall layers and connective abundantly.
- At late sporogenous stage, all wall layers of the anther show rich starch grains. Endothecium and middle wall layers are containing more amounts of PAS +ve.
- At the meiocyte stage, all the wall layers are richly stained with PAS +ve and contain starch grains except tapetum. Meiocyte shows a little number of polysaccharides.
- At the tetrad stage, the tetrahedral tetrads are enveloped by a richly stained callose wall. Starch grains remain same except tapetum.
As development proceeds, the microspores are moderately stained with PAS-positive with the nucleus. Tapetum and two middle wall layers are intact and contain a rich amount of polysaccharides. The starch grains are only present in endothecium Fig. 7. At the vacuolated microspore stage, the microspores are richly stained with insoluble polysaccharides. Initially, the tapetum is single-layered but later, some of the cells undergo the periclinal division so that it becomes two-layered at this stage. The richly cytoplasmic tapetal cells are at first uninucleate but later it undergoes mitotic divisions; they become binucleate and each cell contains 1 or 2 nuclei Fig. 8. The pollen grains are very rich PAS-positive with rich starch storage materials. The endothecium becomes enlarges and the epidermis becomes papillate at this stage. Tapetum is intact and shows vacuoles in it and it is moderately stained with PAS-positive Fig. 9. The endothecium becomes enlarges and fibrous band thickenings are formed, these fibrous thickenings are moderately stained with polysaccharides. Pollen grains are darkly stained with PAS-positive. One middle wall layer degenerates and one is starting to disintegrate and another one is persistent and is moderately stained Fig. 10. The dehiscence takes place through stomium and is longitudinal in nature. Epidermis, endothecium and pollen grains are shows a rich amount of polysaccharides. Tapetum is disintegrated and some traces of tapetum are remaining Fig. 11. The mature anther is completely dehisced and richly stained pollen grains are released. Here one middle layer is still persistent at this stage. The fibrous band thickenings are richly stained with insoluble polysaccharides Fig. 12.
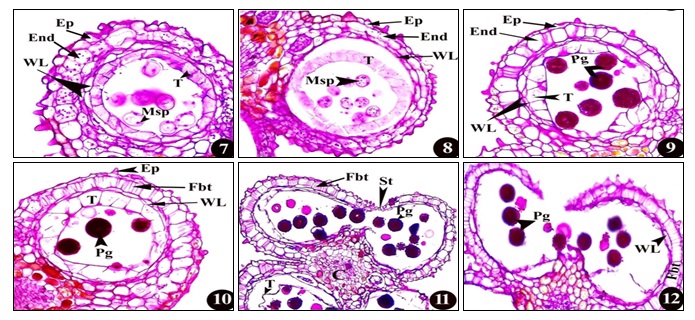
PLATE 2: TRANSVERSE SECTION OF MELIA AZEDARACH L. ANTHERS TESTED FOR INSOLUBLE POLYSACCHARIDES. (EP- Epidermis; End-Endothecium; WL- Wall layers; T- Tapetum; Msp- Microspores; Pg- Pollen grains; C- Connective; FBT- Fibrous band Thickening; St- Stomium;)
- Microspores are moderately stained with PAS +ve with the nucleus. All the wall layers are richly stained with polysaccharides and starch grains are restricted to only endothecium at this stage.
- The epidermis, endothecium, middle wall layers and microspores are shows more amounts of polysaccharides. Tapetum is moderately stained and shows bilayered at this stage.
- The pollen grains contain rich amount of polysaccharides with a filling of starch grains. Endothecium becomes enlarges and tapetum is intact at this stage.
- Epidermis and fibrous band thickenings are moderately stained with PAS +ve. Pollen grains are very rich in PAS +ve. Depletion of starch grains in all the layers.
- At a dehisced stage, epidermis and pollen grains are rich in polysaccharides. Fibrous band thickenings are moderately stained and stomium is seen at this stage. Tapetum starts degeneration.
- The pollen grains are dehisced and tapetum is completely disintegrated. Fibrous band thickenings are moderately stained with PAS +ve. Only one middle wall is persistent.
DISCUSSION: Histochemical methods have been developed for the qualitative and quantitative analysis of virtually all cellular components, including proteins, carbohydrates, lipids, nucleic acids and the range of ionic elements occurring in cell solution 19, 20. Histochemistry combines the techniques of biochemistry and histology in the study of the chemical constitution of cells and tissues. As such, it is concerned with the identification and distribution of various chemical components of tissues through the use of stains, indicators as well as microscopy. Biochemical studies have provided a reasonably complete catalog of the major polysaccharides and phenolic substances that constitute the primary cell walls of angiosperms 21, 22. The anther is one of the principal reproductive organs in the flowering plants. Its structure and cytology have been well documented 23. Histochemical information is available on various aspects of anther development 16, 18, 24-35.
The anthers of Melia azedarach L. is tetrasporangiate. But in Smilax davidiana, the anther is bisporangiate 36. In Cucumis melo individual bisporangiate stamens strongly suggest the double nature of tetrasporngiate stamens 37. Sporogenous tissue forms a central and key tissue for all the vital happenings that occurs in the anther locules. The sporogenous cells generally possess thin cell wall which are PAS-positive suggesting their extensibility, a pre-requisite for rapid cell division and cell expansion. Early sporogenous cells are thin-walled in Dendrobium ovatum and are poor in polysaccharides, as the sporogenous cells multiply in a number them retain rich cytoplasmic polysaccharides and protein content 38. But in Melia azedarach L. the sporogenous cells are moderately stained with PAS-positive.
K. Garudamma in 1957 39 reported that in Melia azedarach L., the anther is four lobes become conspicuous, and at this stage, the sporogenous cells can be distinguished from an outer wall consisting of a tapetal layer. Three middle layers are reported and the tapetal cells remain intact till the tetrads of microspores are formed and the gradually disintegrate. The microspore mother cells undergo meiotic divisions to form microspore tetrads which are usually of the tetrahedral type but occasionally exhibit a decussate arrangement also. The pollen grains are tetracolporate. N. C. Nair in 1959 reported five wall layers in the anthers of Naregamia alata and the secretary tapetum cells to become multinucleate. The reduction division is simultaneous as is also the case in Dysoxylum malabaricum 40, Azadirachta indica 39 and Cipadessa baccifera 41, but in Sandoricum koetjape 42 they are said to successive. The microspore tetrads are tetrahedral or decussate, the tricolporate pollen grains are shed at the three celled stages. Wall formation usually receives little attention in the study of anther ontogeny. The anther wall is formed by a specific number of cell layers that originate in the earliest developmental stages 43. Melia azedarach L. is having one or two middle layers and multinucleate tapetum 12. In the present study Melia azedarach L. anthers show six wall layers i.e. a single layer of epidermis and endothecium, three middle wall layers and a single layer of the tapetum. But the previous study reported that, up to six middle wall layers formed in Normania triphylla and nine in Hawkesiophyton panamense 43. In Ipomea hederifolia and Ipomea macrantha one middle wall layer is seen 44. In Smilax davidiana, the anther is consists of four anther wall layers i.e. one epidermis, one endothecium, one middle wall layer and one tapetal layer. Ricinus communis anther wall consist of a single layer of cells was referred to as an endothecium 45. A multilayered endothecium has been observed in some Solanaceae such as Nicotiana glutinosa and N. tobaccum 46.
In Smilax davidiana the epidermis is uniseriate and persistent which develops fibrous thickenings after the anther wall has ripened 36. As reported by Jain (1956), in Lycium europaeum a multilayered endothecium was observed in the area of connective tissue of the anthers. While 47 Zhukora and Poddubnaya (1987) showed the formation of two-layered endothecium in Lycopersicon esculentum. Maheshwari (1971) 48 who reported 10-14 middle layers in Vallaris foetida. Agadi S. N. and Talawar P. L.(2017) 49 reported basic type of wall formation i.e. single layer of epidermis and endothecium, two middle wall layers and bilayered glandular tapetum in Solanum viarum. The anther wall development in Liliaceae was reported as monocotyledonous type 43, including Gasteraia verrucosa 50, Anemarrhena asphodeloides, Ophiopogon xylorrhizus 51, and Ornithogalum virens 8. But in present study Melia azedarach L. the Basic type of anther wall formation is seen. Talawar P. L. and Agadi S. N. (2018) 16 reported that there is no starch grains present in the anther walls of the Azadirachta indica A. Juss. But in present study Melia azedarach L. has abundant starch grains in all the wall layers of the anther in initial stage. Johnson (1888, 1889), Heinricher (1915), Staedtler (1923), Pisek (1924) and Dowding (1931) 52-56 report that in Arceuthyobium oxycedri the epidermis itself acquires fibrous thickenings and functions as an endothecium. But in the present study, the epidermis and endothecium are formed separately and endothecium enlarges and becomes fibrous thickening at the mature stage of the anther.
According to Pisek (1923) 56 there is no tapetum in Viscum album. In Rhizophora mucronata, the tapetum is a triplet in origin, formed by inner secondary parietal, connective and the lateral sterile cells. (Sudhakaran S. et al., 1995) 57 and in Ipomea pes-caprae, tapetum is dual in origin and dimorphic in nature 44. But in the present study, the secretary type of tapetum is seen. Amoeboid tapetum which was 2-4 layered was seen in Araceae family. It was also seen in Tradescantia L. (Liliaceae), Alisma L. (Alismataceae), Butomus L. (Butomaceae), Typha L. (Typhyaceae) 58. Meiocytes are the ultimate formation of sporogenous cells through differentiation as seen strikingly in their specific cellular activity during meiosis. Many investigators have tried to contribute to the understanding of these changes in meiocytes 26, 49, 59-62. In Melia azedarach L., during the meiocyte stage, we can observe the deposition of the starch grain in all the wall layers except tapetum and they show PAS-positive.
The decussate types of tetrads are reported in Brucea amarissima 63 and isobilateral, decussate tetrads were seen in Samdera indica 64. Decussate tetrads were seen in Tarenna gracilipes 65. Asphodelus aestivus underwent simultaneous cytokinesis in the PMCs and isobilateral microspore tetrads 43. The successive type of cytokinesis is seen in PMCs during meiosis, as in most of the monocotyledons 66 and the other Liliaceae members 43. In Gagea villosa dyads were found as well as tetrads in the same anther locus and the tapetum is begun to degenerate in tetrad stage only 67. But in the present work, the cytokinesis is simultaneous and the tetrahedral tetrads are formed. In Melia azedarach L. Narasimhachar (1936) 68 reported that linear tetrads are present.
Isobilateral and tetrahedral tetrads were seen in Tulipa clusiana 69. The histochemical analysis shows the absence of callose from the walls of the microspore tetrad in Pergularia daemia 70. But in the present study, the tetrahedral tetrads are shows thick callose wall with PAS-positive stain. The role of the callose wall has been implicated in the determination of exine formation 71. In the present investigation refilling of the starch grain in the vacuolated microspores from the young pollen gains. According to Wang and Chen (2001) Oxalis acetocella pollen grains are tetracolporate and rarely tri or pentacolporate. But in present work the pollen grains are tetracolporate. In Solanum viarum, the pollen grain shows degenerating tapetum rich in insoluble polysaccharides 49. But in present investigation the tapetum is intact at pollen grain stage. In Dysoxylum binectariferum Paetow (1931) 40 reports that the shedding occurs at the one-celled stage. In our present work pollen has distinct nuclei at the center with exine and intine wall, the pollen grains and fibrous band thickenings are rich in polysaccharides. One middle wall layer is persisting at dehiscence stage. Stomium is thin-walled and dehiscence takes place through it.
CONCLUSION: The present ontogenically and histochemical study on developing anthers of Melia azedarach L. is an attempt to identify histochemical localization with microsporogenesis and histochemistry reveals the following features.
Melia azedarach L. has 10 anthers. The anther is tetrasporangiate and basic type of wall formation is seen. It has a single epidermis which becomes papillate during the dehiscence stages. Endothecium is single-layered and in mature anther, it enlarges and becomes fibrous bands of thickenings. Three middle wall layers are found in Melia azedarach L. Secretary type of tapetum is reported and which are nutritional in function. The primordium consists of an undifferentiated mass of thin-walled cells and having a uniform distribution of polysaccharides. During the early stages, the rich content of polysaccharides is observed in the wall layers of anthers. Moderate amount of polysaccharides is present in sporogenous, meiocyte and tapetal cells.
Meiocyte shows a moderate amount of polysaccharides with a prominent nucleus. The tetrahedral tetrad is deeply stained with surrounding richly stained thick callose wall. After the dissolution of the callose wall, the individual young microspores are released and later the young microspores become vacuolated microspores. These microspores are rich in polysaccharides and the tapetum becomes two-layered at this stage. Tapetum is intact up to the pollen grain stage. The pollen grains contain a high amount of insoluble polysaccharides with a filling of abundant starch grains. The multiple fibrous band thickening is formed in the dehisced stage. Finally, it is concluded that there are interaction and utilization of biochemical substances in the anthers during the formation and differentiation of pollen grains. Anther development at all the stages is normal, synthesis and lyses of the macromolecular biochemical observed at specific stages of the anther development. It shows pollen may be viable and germinate after pollination.
ACKNOWLEDGEMENT: Authors acknowledge the Guide Dr. S. N. Agadi and Chairman, P.G. Department of Botany, Karnatak University, Dharwad for extending facilities. Also, acknowledge the financial assistance from Rajiv Gandhi National Fellowship to the author.
CONFLICTS OF INTEREST: Nil
REFERENCES:
- Ma H: Molecular genetic analyses of microsporogenesis and microgametogenesis in flowering plants. Annual Review of Plant Biology 2005: 56, 393-34.
- Sun YJ, Hord CLH, Chen CB and Ma Hc: Regulation of Arabidopsis early anther development by putative cell-cell signaling molecules and transcriptional regulators. Journal of Integrative Plant Biology 2007: 49: 60-68
- Li, Adams L and Chen S: Eugenia jambolana Berry extract inhibits growth and induces apoptosis of human breast cancer but not non-tumorigenic breast cells. Journal of Agricultural Agric Food Chem 2009; 57(3): 826-31.
- McCormick S: Control of male gametophyte development. Plant Cell 2004; 16: S142-S153.
- Pacini E and Franchi GG: Types of pollen dispersal units in monocots. Austr J Bot (in press) 2000.
- Pacini E and Hesse M: Pollenkitt - its composition, forms and functions. Flora - Morphology, Distribution, Functional Ecology of Plants 2005; 200(5): 399-15.
- Bohdanowicz J, Szczuka E, Świerczyńska J, Sobieska J and Kościńska-Pajak M: Distribution of microtubules during regular and disturbed microsporogenesis and pollen grain development in Gagea lutea (L.) Ker.-Gaw. Acta Biologica Cracoviensia 2005; 47: 89-96.
- Lesniewska J and Charzynska M: Tapetal plastids in Ornithogalum virens: from meristematic stage to pollen coat. Acta Biologica Cracoviensia 2000; 42: 141-49.
- Taylor ML and Osborn JM: Pollen ontogeny in Brasenia (Cabombaceae, Nymphaeales). American Journal of Botany 2006: 93: 344-56.
- Johri BM, Ambegaokar KB and Srivastava PS: Comparative embryology of angiosperms. Berlin: Springer-Verlag 1992.
- Nair NC: Studies on Meliaceae 2. Floral morphology and embryology of Melia azedarach: A reinvestigation. J Indian Bot Soc 1959; 38: 367-78.
- Galati BG, Sonia R and Monica TG: Embryological studies in Melia azedarach J Tor Bot Soc 2011; 138(1): 26-33.
- Chang KT and Wang FH: Morphology of pollen grains of Meliaceae. [In Chinese: English summary.]- Pollen morphology. Meliaceae. Act Bot Sinica 1956: 5: 253-65.
- Garudamma GK: Studies in Meliaceae 1.Development of the embryo in Azadirachta indica. A. Juss. J Ind Bot Soc 1956: 35: 222-26.
- Dida S, Endah RP, Iskandar ZS and Andry I: Flower initiation, morphology and developmental stages of flowering-fruiting of Mindi (Melia azedarach) JMHT 2012.
- Talawar PL and Agadi SN: Anatomy and distribution of polysaccharide during anther development in Azadirachta indica JUSS. J of Ind Bot Soc 2018; 97(1): 12-20.
- Feder N and O’Brien TP: Plant microtechnique: some principles and new methods. Amer J Bot 1968; 55123-42.
- Panchaksharappa MG, Rudramuniyappa CK and Hegde RR: A review on recent advances in histochemistry of anther development. J Pl Sci Res 1985; 1: 60-83.
- Gahan PB: Plant Histochemistry and Cytochemistry, Academic Press, London 1984.
- Kiernan JA: Histological and histochemical method. Theory and Practice. 3rd Oxford UK: Butterworth- Heinemann. 1999.
- McCann MC, Shi J, Roberts K and Carpita NC: Changes in pectin structure and localization during growth of unadapted and NaCl-adapted tobacco cells. The Plant Journals 1994; 773-85.
- Carpita NC and Gibeaut DM: Structural models of primary cell walls in flowering plants: consistency of molecular structure with the physical properties of the walls during growth. The Plant Journal 1993; 3: 1-30.
- Maheshwari P: An Introduction to the Embryology of Angiosperms. Tata McGraw-Hill, New York 1950.
- Heslop-Harrison J: Pattern in plant cell walls: Morphogenesis in miniature. Proc R Inst Great Britain 1972; 4(5): 335-51.
- Mascarenhas JP: The biochemistry of angiosperm pollen development. Bot Rev 1975; 41: 259-14.
- Bhandari NN and Sharma M: Ontogenetic and histochemical studies on the anther of Carthamus tinctorius Jour Palynol 1983; 19: 153-80.
- Blackman SJ and Yeung EC: Comparative anatomy of pollinia and caudicles of an orchid (Epldendrum). Bot Gaz 1983; 144: 331-37.
- Shivanna KR and Johri BM: The angiosperm pollen: structure and function. Wiley Eastern Ltd., New Delhi. 1985; 53-83.
- Hegde RR and Issac SW: Cytology and histochemistry of microsporogenesis in cytoplasmic male sterile plants. In: C.P. Malik (Ed.). Advances in pollen-spores research: XIX, Pollen physiology and biotechnology. Today and Tomorrows Printers and Publishers, New Delhi 1992: 41-63.
- Hegde RR, Halappanavar SP and Agadi SN: Morphogenetic events during anther development. In: XIX, C.P. Malik (Ed.) Advances in pollen-spore Research; New perspectives in pollen spore Biology 1993: 1-36.
- Katti RY, Giddanavar HS, Shamala N, Agadi SN and Hegde RR: Persistance of callose and tapetum in the microsporogenesis of genic male sterile Cajanus cajan, (L).Millsp with well formed endothecium .Cytologia 1994; 59: 65-72.
- Agadi SN: A comparative cyto-histochemical observations of microsporogenesis in male sterile and male fertile rice. Ph. D. Thesis Karnatak University, Dharwad 1996.
- Jayaraj M: Embryological of Penstemon nitidus Dougl. (Cheloneae-Scrophulariaceae) Jour Kar Uni Dharwad 2003; 60-67.
- Agadi SN and Negalur CB: Ontogeny and Histochemistry of anther development in Russelia equisetiformis. Schlecht and Chan Acta Bio Ind 2014; 3(1): 494-98.
- Agadi SN and Naik V: Ontogeny and Histochemistry of polyad of Dendrobium ovatum (wild). Kranz. Acta Bio Ind 2012; 1(2): 207-13.
- Cheng-Qi A: Developmental origins of the conjoined twin mature embryo sacs in Smilax davidiana, with special notes on the formation of their embryos and endosperms. Amer J Bot 2013: 100(12): 2509-15.
- McLean DM: Stamen Morphology in flowers of the Muskmelon. J of Agricultural Res 1947; 74(2): 49-53.
- Agadi SN and Naik V: Ontogeny and Histochemistry of Polyad of Dendrobium ovatum (wild). Kranz Acta Bio Ind 2012; 1(2): 207-13.
- Garudamma GK: Studies in Meliaceae 1. Gametogenesis in Melia azedarach. L. J Ind Bot Soc 1957; 36: 227-31.
- Paetow W: Embryologische Untersuchungen an Tacca-ceen, Meliaceen und Dilleniaceen. Pla 1931; 14: 441-70.
- Narayana LL: Floral anatomy and embryology of baccifera Miq. J of the Ind Bot Soc 1958a; 37: 147-54.
- Juliano JB: Studies on the morphology of the meliaceae. II. Sterility in santol, Sandoricum koetjape (Burm. f.) Merr. Phil 1934; 253-62.
- Davis GL: Systematic embryology of Angiosperms- J. Wiley and Sons, New York 1966.
- Lakshminarayana K and Rao PS: Embryological investigations in some species of Ipomea. Taiwania 1993; 38: 109-16.
- Van der Pijl L: The stamens of Ricinus. Phytomorphology 1952; 2: 130-32.
- Jose J and Singh SP: Gametophyte development and embr-yogeny in the genus Nicotiana. JIBS 1968; 47: 117-28.
- Zhukora and Poddubnaya A: AVA Solanaceae,- In: Batygina, T. B and Yakovlev, M.s. (eds). Comparative Embryology of Flowering Plants 1987; 4: 241-47.
- Maheshwari DH: Embryology of Apocynaceae. I. Plumiereae. J Indian Academy of Science 1971; 60: 52-65.
- Agadi SN, Talawar PL and Emmi SN: Ontogeny of anther in viarum Dunal. Int J Appl Res 2017; 2(6): 86-91.
- Keijzer CJ: The process of anther dehiscence and pollen dispersal. I. The opening mechanism of longitudinally dehiscing anthers. New Phytologist 987; 105: 487-98.
- He TH, Rao GY, You RL and Zhang DM: Embryological studies on endangered Ophiopogon xylorrhizus. Acta Phytotaxonomica Sinica 1998b; 36: 305-09.
- Johnson T: Arceuthobium oxycedri. Ann Bot 1888; 2(2): 137-60.
- Johnson T: The nursing of the embryo and some points in Myzoderidron punctulatum Bankset Sol. Ann. Bot., Lond 1889; 3: 179-06.
- Heinricher H, Bau H und Biologie der Bliiten von Arceuthobium Ozycedri. S.B. Akad Wiss Wien 1915: 124: 181-230.
- Staedtler G: Über Reduktionserscheinungen im Bau der Antheren wand von Angiospermen-Blüten. Flora 1923; 116: 85-08.
- Pisek A: Chro1nosomenverh~1tnisse, Reduktionsteilung und Revision der Keimentwicklung der Mistel (Viscum album). Jb. wiss Bot 1923; 62: 1-19.
- Sudhakaran S, Vaidyanathan R and Ganapathi A: Microsporogenesis in a mangrove plant Rhizophora mucronata - Thaiszia -J. Bot Košice 1995; 5: 27-30.
- Ünal M: Plant (Angiosperm) Embryology. ISBN 975-400-040-9, İstanbul, TURKEY 2004.
- Dickinson HG and Potter U: Post-meiotic nucleocytoplasmic interaction in Cosmos bipinnatus: Early events at the nuclear envelope. Planta 1979; 145: 449-57.
- Dickinson HG and Heslop-Harrison J: Ribosomes, membranes and organelles during meiosis in angiosperms. Phil Trans R Soc Lond B 1977; 277: 327-42.
- Vijayaraghavan MR and Bhatia K: Cellular changes during microsporogenesis, vegetative and generative cell formation; a review based on ultrastructure and histochemistry. Int Rev Cytol 1985: 96: 263-96.
- Dickinson HG: The physiology and biochemistry of meiosis in the anther. Int Rev Cytol 1987; 107: 79-09.
- Nair NC and Sukumaran NP: Floral morphology and Embryology of Brucea amarissima. Botanical Gazette 1960; 121: 175-85.
- Nair NC and Joseph TC: Floral morphology and Embryology of Samdera indica. Botanical Gazette 1957; 119: 104-15.
- Vinckier S and Smets E: A histological study of microsporogenesis in Tarenna gracilipes (Rubiaceae). Grana 2005; 44: 30-44.
- Furness CA and Rudall PJ: Microsporogenesis in monocotyledons. Ann of Bot 1999; 84: 475-99.
- Nuran E: Microsporogenesis, pollen mitosis and pollen tube growth in Gagea villosa (Liliaceae). Bio 2014; 69(10): 1323-30.
- Narasimhachar SG: Megasporogenesis and embryo sac formation in two swpecies of meliaceae. Current Science 1936; 298-99.
- Wafai BA and Koul AK: Analysis of breeding systems in Tulipa. II. Sporogenesis, gametogenesis and embryogeny in tetraploid T. clusiana. Phytomorphology 1982; 32(4): 289-01.
- Vijayaraghavan MR and Shukla AK: The Nature of Covering around the Aggregate of Microspores in Pergularia daemia (Forsk.) McC & Blat Ann Bot 1976; 40: 417-21.
- Waterkeyn L and Bienfait: On a possible function of the callosic special wall in Ipomea purpurea (L.) Roth 1970: 10(1): 13-20.
How to cite this article:
Talawar PL and Agadi SN: Ultrastructure and histochemistry of anther development in Melia azedarach L. Int J Life Sci & Rev 2019; 5(11): 162-71. doi: 10.13040/IJPSR.0975-8232.IJLSR.5(11).162-71.
All © 2015 are reserved by the International Journal of Life Sciences and Review. This Journal licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 3.0 Unported License.
Article Information
2
162-171
997
1203
English
IJLSR
P. L. Talawar * and S. N. Agadi
Developmental Biology and Histochemistry Laboratory, P. G. Department of Botany, Karnatak University, Dharwad, Karnataka, India.
talawarpoornima@yahoo.com
21 October 2019
21 November 2019
26 November 2019
10.13040/IJPSR.0975-8232.IJLSR.5(11).162-71
30 November 2019